| 図 1 リボソーム小サブユニットRNAの比較に基づく生物の系統樹。生物界は3つのグループからなる。真核生物(Eukaryoteまたは Eukarya)、真正細菌(EubacteriaまたはBacteria)、古細菌(ArchaebacteriaまたはArchaea)。太い線は超 好熱菌(至適生育温度80℃以上の菌)をむすんでいる。系統樹の根本の点線の下にある「起源生命」は、last universal common ancestor (LUCA) などとも呼ばれるが、RNAの比較からではなく蛋白質のパラログの比較などから、この「根」の位置が想定される。この位置は超好熱菌に囲まれているため、最初の原始生命は海底熱水噴出口のような超好熱性の環境で誕生したと推測される)。 |
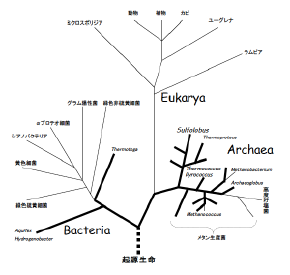
1977 年Woeseは、リボソーム小サブユニットRNAの塩基配列の比較から、生物を、真核生物(Eukaryote)、真正細菌(Eubacteria)、古 細菌(Archaebacteria)の3つのグループに分類することを提唱した。その後の比較生化学的研究から、古細菌は、真正細菌と類似する性質と、 蛋白質合成系の一部にみられるような真核生物との類似性と、エーテル型脂質等に見られる古細菌独自の性質とを併せ持ち、生物の進化系統上始原的な位置を占 めると考えられるようになり、意を強くしたWoeseはArchaebacteriaという名称をArchaeaと呼び変えるようになった[図1]。 今日分離されている多数の古細菌の中には、真正細菌とは異なる原始的と思われる代謝系や酵素蛋白質をもち、真核生物の祖先型代謝を太古から維持していると 考えられるものがある。古細菌は、絶対嫌気条件や飽和食塩水や超高温のような、他の普通の生物の生存できない極限環境を好むものが多く、こうした厳しい環 境の中で、のんびりとがんばっている。
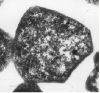 |
| 図2 好酸好熱性古細菌 Sulfolobus tokodaii (pH 2.5 ,75℃で培養)の切片写真。Sulfolobusは不規則形の古細菌である。 |
リボソーム小サブユニットRNAの塩基配列などの比較から、地球上に最初に発生した生物は1種類であり高度に好熱性であったと考えられる。生育至適温度が80℃をこえるものは超好熱菌と呼ばれ、105℃を至適とするものもある。超好熱菌の大部分は古細菌である[図1]。超好熱菌の生体高分子は、熱だけでなく物理化学的に安定で、その安定性の解析や応用の視点から注目される。さらに、超好熱菌には、特異な代謝系が見られることが多く、それに関わる特異な酵素を有するという点からも興味深い生物である。
当研究室では、超好熱菌に特有の糖/エネルギー代謝経路の酵素を主な対象として、蛋白質の構造と機能の相関を調べ、生命の進化・生体高分子の理解や、新たな応用に資することを目指している。
 |
|
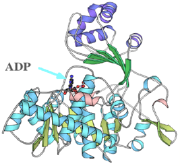
図3 超好熱性古細菌Thermococcus由来ADP依存性グルコキナーゼの立体構造。普通の生物がATPを用いてグルコースをリン酸化するのと異なって、ADPをリン酸供与体とする酵素で、解糖系の初段で働いている。
|
これまでに、我々が研究に取り組んできた特異な酵素の例を挙げると、次のようなものがある。
1)4-α-グルカノトランスフェラーゼ(直鎖アミロースからの巨大環状糖の形成)
2)ADP依存性グルコキナーゼ、ADP依存性ホスホフルクトキナーゼ(ADPでなくATPをリン酸供与体とする解糖系酵素。その立体構造を初めて明らかにした。超好熱菌の一部しか持っていないと思われていたが、マウスやヒトにも類似酵素があることが最近わかった)[図3]。
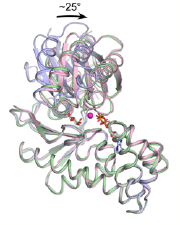 |
| 図4 好酸好熱性古細菌S. tokodaii 由来ヘキソキナーゼが、糖基質の結合によって大きな構造変化を起こす様子。結合の前後の基本骨格を重ね描きしてある。この酵素は、ATPを用いるものの、ふつうのヘキソキナーゼと全く異なるアミノ酸配列を示す特異な酵素で、糖新生系で働くと思われる。詳しくは、こちらのリンク、動画はこちらのリンクを参照。 |
3)ATP依存性ヘキソキナーゼ(普通は解糖系初段の酵素だが、ある種の超好熱菌ではアミノ酸配列が全く異なり解糖系でなく糖新生系で働くものがある。これを結晶化して構造を調べ、これまで分からなかった、マグネシウムの働きや大きな構造変化を明らかにした)[図4]。
4)鉄を含むクピン型のホスホグルコースイソメラーゼ(PGI、解糖系の酵素だが、普通のPGIとはアミノ酸配列が全く異なる。その立体構造を初めて明らかにした)
5)フルクトースビスホスファターゼ(FBPase、糖新生系で働く酵素で、古細菌の酵素は他の生物の酵素とアミノ酸配列が全く異なる「古細菌型」。その立体構造を初めて明らかにした。)
6) 2-オキソ酸:フェレドキシン酸化還元酵素(NADでなくフェレドキシンを電子受容体とするクエン酸回路入り口及び途中の酵素で、分子量20数万以下の簡 素な構造をしている。普通の生物は分子量数百万の複雑な酵素を使っている)及びフェレドキシン(フェレドキシンは鉄硫黄クラスターを活性中心とする酸化還 元蛋白質であるが、超好熱菌の中には鉄に加えて亜鉛を持つもの[図5]がある。)
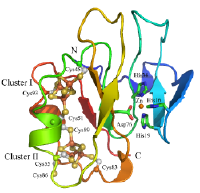 |
| 図5 S. tokodaii 由来フェレドキシンの立体構造。N-末端の37残基の付加配列が小さいドメイン(図の右側)を、残りの66残基が二つの鉄硫黄クラスターを含むコアドメイン(図の左側)を形成し、両ドメインの界面に亜鉛が結合して、分子を安定化している。 |
7)好気的な呼吸鎖電子伝達系の成分 (高温で希薄な酸素を利用する呼吸系)
8)細胞質膜酵素や細胞表層蛋白質の研究
9)硫黄の酸化還元に関わる巨大酵素の立体構造と機能の解析
これらの中には、生物の重要な代謝経路である解糖系(やその逆の糖新生系)に関わるものが多いが、超好熱菌の中には、普通の生物とは異なる解糖系路を持つ ものがあり、それを触媒する酵素についても明らかでない場合がある。超好熱菌のゲノム解析が進んでも遺伝子と対応のつかない酵素や蛋白質は多い。2. その他の有用酵素の構造と機能の解析
超好熱菌の他にも、産業上利用価値の高いと思われる酵素(例えばニトリルヒドラターゼ)の構造や機能の解析を行っている。また、進化的にATPの先駆と考 えられるPPi(無機ピロリン酸)に依存する酵素(例えば超好熱菌の無機ピロホスファターゼ)についても研究を進めている。
連絡先
E-mail: t-wakagi[at-mark]cb3.so-net.ne.jp